| Физиологические основы траспорта в крови жирных кислот |
Для
переноса в крови (водной среде)
гидрофобных, т. е. водонерастворимых веществ,
сформированы транспортные системы.
Несмотря на существенное различие
гидрофобных веществ (кислород, билирубин, стероиды, тиреоидные гормоны,
жирорастворимые витамины, ионы
двухвалентных металлов), транспорт
их к клеткам основан на единых
принципах.
1. Основу каждой транспортной
системы составляет транспортный
белок.
2. Все транспортные белки переносят
в крови простые вещества.
3. Каждая транспортная система
специфична и переносит к клеткам
одно простое вещество.
Система липопротеинов (ЛП) переносит в кровотоке жирные кислоты (ЖК). При этом аполипопротеином (апо) является белок, который формирует белок-липидный комплекс (БЛК) из одного класса липидов, определяет функциональные свойства БЛК и вызывает гиперлипопротеинемию (ГЛП) при генетическом дефекте или отсутствии синтеза апобелка.
Структурно ЖК подразделяют на насыщенные и ненасыщенные (Табл. 1). Насыщенные кислоты и мононенасыщенная олеиновая кислота, окисляясь в митохондриях, поставляют клетке основное количество АТФ. Полиненасыщенные или полиеновые ЖК выполняют структурную функцию, определяют специфичность клеточных мембран высокодифференцированных клеток и являются предшественниками синтеза эйкозаноидов (простагландины, тромбоксаны, лейкотриены).
Физико-химическая индивидуальность ЖК настолько велика, что транспортировать разные свободные ЖК в одном ЛП невозможно. Индивидуальность ЖК можно снизить, если ковалентно связать несколько ЖК с одной матрицей. В качестве матрицы служат спирты с разной степенью гидрофобности (глицерол, холестерол, сфингозин). Однако, и после этого насыщенные и ненасыщенные ЖК оказываются в разных сложных липидах. Триглицериды являются транспортной формой преимущественно насыщенных, а фосфолипиды - преимущественно ненасыщенных ЖК.
Табл. 1 Жирные кислоты эукариот.
| Атомы
углерода: число и положение двойных связей |
Семейство | Название | Систематическое наименование | Распространение |
| Насыщение | ||||
| 16:0 | - | Пальмитиновая | Гексодеканоевая | Все растительные и животные жиры |
| 18:0 | - | Стеариновая | Оптадеканоевая | То же |
| Моноевая | ||||
| 16:1;9 | w-7 | Пальмитоолеиновая | цис-Гексадеценовая | " |
| 18:1;9 | w-9 | Олеиновая | цис-Октадеценовая | " |
| 18:1;9 | w-9 | Эллаидиновая | транс-Октадеценовая | Жиры жвачных, процесс гидрогенизации |
| 22:1;13 | w-9 | Эруковая | цис-13-Докозеновая | Рапсовое и горчичное масло |
| Диеновая | ||||
| 18:2;9,12 | w-6 | Линолевая | цис-9.12-Октадекадиеновая | Растительные масла |
| Триеновая | ||||
| 18:3;6,9,12 | w-6 | g-Линоленовая | цис-6,9,12- Октадекатриеновая | Льняное масло |
| 18:3;9,12,15 | w-3 | а-Линоленовая | цис-9,12,15- Октадекатриеновая | То же |
| Тетраеновые | ||||
| 20:4;5,8,11,14 | w-6 | Арахиденовая | цис-5,8,11,14- Эйкозотетраеновая | Арахисовое масло |
| Пантаеновые | ||||
| 20:5;5,8,11,14,17 | w-3 | Тиинодоновая | цис-5,8,11,14,17- Эйкозопентаеновая | Рыбий жир |
| 20:5;7,10,13,16,19 | w-3 | Клупанодоновая | цис-7,10,13,16,19- Докозопентаеновая | Рыбий жир, фосфолипиды ткани мозга |
| Гексаеновые | ||||
| 22:6;4,7,10,13,16,19 | w-3 | Цервоновая | цис-4,7,10,13,16,19- Докозогексаеновая | То же |
Липидами являются ЖК и все сложные липиды-производные Ж.К (триглицериды, фосфолипиды, гликолипиды, цереброзиды, ганглиозиды). Холестерин является спиртом, но после взаимодействия с ЖК образованные эфиры холестерина являются сложными липидами.
Простые и сложные липиды делятся на полярные (свободные ЖК, моно- и диглицериды, фосфолипиды; полярным является и холестерин). К неполярным липидам относятся триглицериды и эфиры холестерина. Полярные липиды формируют монослойные структуры. Неполярные липиды образуют гидрофобную массу. Между неполярными липидами и водной средой всегда формируется монослой полярных липидов.
Табл.2 Перенос простых и сложных липидов транспортными белками.
| Липиды | Транспортный белок 1 | |
| Свободные насыщенные ЖК | Альбумин 1 | |
| Свободные ненасыщенные ЖК | Альфа-фетопротеин | |
| Холестерин | АпоА-1У | |
| Моно-, диглицериды | БПЭХ | |
| Фосфолипиды | АпоА-1 - | |
| Триглицериды | Апо-В-48, апоВ-100 третий домен : | |
| Эфиры холестерина | Апо-В-100 пятый домен : | |
Стационарными апобелками, которые формируют ЛП, являются апоА-1, изобелки апоВ-48 и апоВ-100. Транспортную функцию выполняют также белок, переносящий эфиры холестерина (БПЭХ), альбумин и альфа-фетопротеин (Табл. 2). В превращении в крови полярных липидов в неполярные задействованы функциональные апоЛП (апоА-11, апоС-11 и апоС-111) и ферменты - постгепариновая липопротеидлипаза и печеночная глицеролгидролаза. В свою очередь, в превращении в крови полярных липидов в неполярные участвуют апоА-1, апоА-11, апоА-1У и лецитин-холестерин ацилтрансфераза. Динамичным апобелком с уникальной функцией лиганда является апоЕ. Филогенетически ранними являются ЛП высокой плотности (ЛПВП) и перенос ЖК в полярных липидах. Более поздним и совершенным является транспорт ЖК в неполярных липидах. Каждый апобелок формирует БЛК из одного класса простых или сложных липидов. АпоА-1 связывает только полярные фосфолипиды. АпоВ-100, имея два разных липидсвязывающих домена, формирует два БЛК - один из триглицеридов, второй из эфиров холестерина (Табл. 3). Несмотря на название, БПЭХ связывает только моно- и диацилглицериды.
Табл. 3. Функция белок-липидных комплексов в транспорте жирных кислот.
| Белок- липидные комплексы |
Функция |
| АпоА-1 | акцептор полярных фосфолипидов |
| АпоА-11 | кофактор лецитин-холестерин ацилтрансфераэы (динамичный БЛК) |
| АпоА-1У | акцептор холестерина (динамичный БЛК) |
| АпоВ-48 | акцептор неполярных триглицеридов (стационарный БЛК) |
| АпоВ-100 | акцептор
триглицеридов третьим и эфиров
холестерина пятым доменом, лиганд для апоВ-100 рецепторов (стационарный БЛК} |
| АпоС-11 | кофактор постгепариновой липопротеидлипазы (динамичный БЛК) |
| АпоС-111 | кофактор печеночной глицеролгидролазы (динамичный БЛК) |
| АпоЕ | лиганд
для апоЕ рецепторов (апоЕ/А-1, апоЕ/В-48 и апоЕ/В-100) (динамичный БЛК |
В крови циркулируют как БЛК, так и ЛП. БЛК являются первичные хиломикроны (ХМ), апоВ-48 БЛК; первичные ЛП очень низкой плотности (ЛПОНП), апоВ-100 БЛК из триглицеридов; ЛП низкой плотности (ЛПНП) апоВ-100 БЛК из эфиров холестерина; ЛП высокой плотности 2а (ЛПВП-2а, апоА-1 БЛК). В ассоциации БЛК формируют ЛП, в которых проходят биохимические превращения липидов (Табл. 4). Липопротеинами являются зрелые ХМ, зрелые ЛПОНП, ЛП промежуточной плотности (ЛППП), ЛПВП-26, ЛПВП-З, ЛП очень высокой плотности (ЛПОВП). Каждый БЛК в ЛП выполняет специфичную функцию. По окончанию биохимических превращений липидов ЛП диссоциируют на отдельные БЛК.
В энтероцитах синтезированные фосфолипиды связывает апоА-1 и формирует первичные ЛПВП. ЛПВП в фосфолипидах переносят ко всем клеткам полиеновые ЖК. АпоА-1 связывается с белками мембраны клеток и при действии фосфолипаз проходит транеэтерификация ЖК: более полиеновая ЖК из фосфолипидов ЛПВП переходит в фосфолипиды мембраны, а менее ненасыщенная ЖК - в обратном направлении; поглощения ЛПВП при этом не происходит. Это - пассивный транспорт в клетки полиеновых омега-6 ЖК. Более длинные и более ненасыщенные омега-З Ж.К так в клетки поступать не могут. В энтероцитах триглицериды связывает апоВ-48 и формирует первичные ХМ, В крови первичные ХМ ассоциируются с апоЕ БЛК и формируют зрелые ХМ. Последние переносят насыщенные ЖК. только в гепатоциты, которые поглощают ХМ путем апоЕ/В-48 рецепторного эндоцитоза. Врожденное нарушение структуры апоЕ и апоЕ/В-48 эндоцитоза приводит к ГЛП типа III.
Функциональное отличие апоА-1 ЛП от апоВ ЛП (ХМ, ЛПОНП, ЛППП и ЛПНП) состоит в том, что первые переносят ЖК только в полярных (простых и сложных) липидах, а вторые - в неполярных сложных липидах. Поэтому, когда в ходе транспорта в апоВ ЛП образуются полярные (простые или сложные) липиды, они переходят в ЛПВП. Когда же в ЛПВП синтезируются неполярные сложные липиды, они переходят в более гидрофобные ЛПНП или ЛПОВП.
ХМ транспортируют в гепатоциты экзогенные насыщенные ЖК с длиной 4-24 атомов углерода. Гепатоциты оптимизируют .ЖК по длине и в секретруемых ими триглицеридах содержатся только С 1биС 18 ЖК. АпоВ-100 связывает триглицериды и формирует первичные ЛПОНП, которые гепатоциты секретируют в кровь. В крови первичные ЛПОНП ассоциируются с апоЕ БЛК и формируют зрелые ЛПОНП. Постгепариновая липопротеидлипаза гидролизует часть триглицеридов ЛПОНП и образуемые полярные свободные ЖК, моно- и диацилглицериды покидают ЛПОНП; насыщенные ЖК связывает альбумин, а моно- и диацилглицериды переходят в ЛПВП-2а, превращаясь при этом в глицерид-содержащие ЛПВП-26. Акцептором моно-, диглицеридов является БПЭХ, В ходе липолиза ЛПОНП превращаются в ЛППП. При генетическом дефекте постгепариновой липопротеидлипазы и нарушении гидролиза триглицеридов в ХМ и ЛПОНП, форми-руется ГЛП типа 1.
Тобл. 4. ЛП кок ассоциоты бепок-липидных комплексов (БЛК)
| ЛП низкой плотности | Ассоциированные БЛК | ЛП высокой плотности | Ассоциированные БЛК |
| Хиломикроны | B-48 | ЛП высокой плотности | A-I |
| Хиломикроны | В-48; Е; С-II; А-I; А-II; A-IV |
пре ЛП высокой плотности | A-I |
| ЛП очень низкой плотности | B-100 | ЛП высокой плотности-1 | A-I; E |
| ЛП очень
низкой плотности |
В-100; Е; С-II; А-II; A-IV |
ЛП высокой плотности- 2а | A-I; (БЛЭХ) |
| ЛП промежуточной плотности |
В-100; Е; С-III; А-II |
ЛП высокой плотности- 2б | A-I; C-III; (БЛЭХ) |
| ЛП низкой плотности | В-100 | ЛП высокой плотности-3 | A-I; A-II; A-IV |
Поскольку ЛППП являются ассоцитатом апоВ-100 и апоЕ БЛК, дальнейший их транспорт может быть разным. При дефиците насыщенных ЖК клетки синтезируют апоЕ, который инициирует поток в клетки насыщенных ЖК в состав ЛППП. Клетки поглощают ЛППП путем апоЕ/В-100 эндоцитоза. Так происходит активный транспорт в клетки насыщенных ЖК. При врожденном изменении структуры апоЕ и нарушении апоЕ/В-100 лиганд-рецепторного взаимодействия формируется ГЛП типа III.
По второму пути липолиз в ЛППП продолжается и освобождаемые свободные С 1биС 18 ЖК связывает альбумин. Альбумин переносит ЖК к миоцитам и адипоцитам, взаимодействует с мембраной клеток, ЖК переходят на мембрану и далее путем диффузии поступают в клетки. Альбумин не связывает полиеновые ЖК, поэтому альбумин формирует пассивны) транспорт в клетки только насыщенных ЖК.
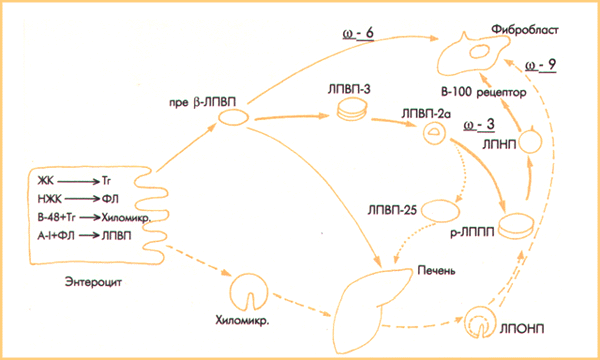
При раздельном транспорте в клетки насыщенных и полиеновых Ж.К и те, и другие поступают в клетки как пассивным, так и активным путем (Рис. 1). Существенное отличие состоит в том, что пассивно в клетки поступают ЖК из полярных липидов без поглощения клетками ЛП. При активном транспорте ЖК поступают в клетку в составе неполярных липидов и при этом клетки поглощают ЛП посредством рецепторного эндоцитоза. Пассивный путь филогенетически более древний; активный транспорт более молодой и более совершенный.
Рис.
1.
Транспорт липопротеинами в крови
насыщенных и полиеновых ЖК.
Сплошные
линии - транспорт
полиеновых ЖК (толстая линия -
активный, тонкая - пассивный).
Точечная линия -
транспорт
полиеновых ЖК в эфирах холестерина.
Пунктирная линия -
транспорт насыщенных ЖК.
Пассивным путем насыщенные ЖК поступают в клетки в связи с альбумином; активным путем насыщенные ЖК поступают в клетки при апоЕ/В-100 рецепторном эндоцитозе ЛППП. Пассивно транспорт омега-6 полиеновых ЖК проходит при транеэтерификации ЖК между фосфолипидами ЛПВП и фосфолипидами мембраны. Однако, омега-З ЖК поступать в клетки пассивным путем не могут. Как же омега-З ЖК поступают в клетки?
Потребность в омега-З ЖК продиктована тем, что клетки рыхлой соединительной ткани используют их для синтеза эйкозаноидов (простагландины, тромбоксаны, лейкотриены). Эйкозаноиды локально регулируют кровоток в капиллярах, агрегации тромбоцитов, экскрецию электролитов, реакцию воспаления. Эйкозаноиды, синтезированные клеткой из арахидоновой омега- 6 и эйкозапентаеновой омега-З ЖК, существенно различаются по активности, Эйкозаноиды действуют всего несколько мин.; депонировать их клетки не могут, поэтому их синтез и действие происходят одновременно. Это требует рецепторного поступления в клетки омега-З ЖК. Однако, рецепторным эндоцитозом клетки поступают только неполярные сложные липиды.
Первым этапом активного транспорта в клетки омега-З ЖК является транеэтерификация ЖК из полярных фосфолипидов в неполярные эфиры холестерина (ЭХ); реакция проходит в ЛПВП. На втором этапе ЭХ переходят в ЛППП при действии БПЭХ. Замещая триглицериды, более гидрофобные ЭХ инициируют превращение ЛППП в ЛПНП. Если ЛПОНП и ЛППП транспортируют в клетки насыщенные ЖК, то ЛПНП переносит только полиеновые, преимущественно омега-З ЖК. Третий этап - поглощение клетками ЛПНП путем апоВ-100 эндоцитоза. Мы полагаем, что апоВ-100 эндоцитоз является ключевым этапом поступления в клетки омега-З ЖК. Генетический дефект структуры апоВ-100 лиганда или отсутствие на мембране апоВ-100 рецепторов приводят к ГЛП типа IIа. Функциональный дефект апоВ-100 лиганда, вызванный неполным замещением триглицеридов на эфиры холестерина и нарушением превращения ЛППП в ЛПНП, составляет основу ГЛП II б типа (Рис. 2).
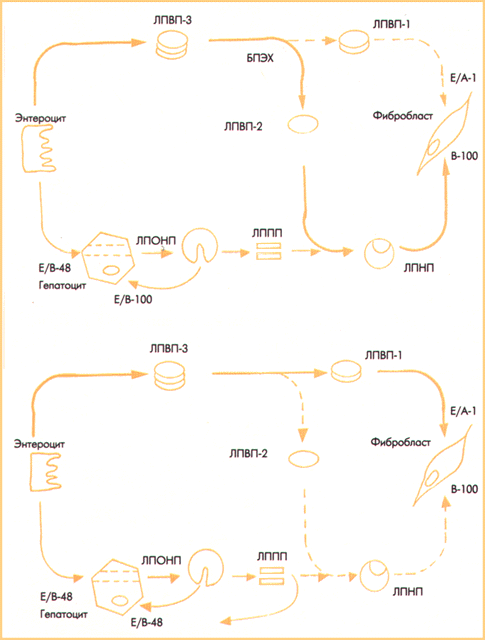
Рис. 2. Кругооборот холестерина в транспорте в клетки полиеновых ЖК и места блокады транспорта ЖК с формированием ГЛП разных типов по Д. Фредриксону.
В лизосомах гидролиз ЭХ освобождает полиеновые ЖК, которые использует клетка. Холестерин же транспортные белки цитозоля переносят на мембрану, с которой он диффундирует в кровь. В крови холестерин связывает апоА-1У и в ЛПВП его опять используют для этерификации следующей полиеновой ЖК. Так происходит кругооборот холестерина в транспорте полиеновых ЖК. Определяя в крови содержание холестерина, мы на самом деле оцениваем уровень ЭХ, которые являются транспортной формой полиеновых ЖК и которые клетки не могут поглотить. Следовательно, холестерин крови отражает дефицит в клетках полиеновых ЖК.
Однако, апоВ-100 рецептор является ключевым этапом транспорта полиеновых ЖК только у части видов млекопитающих (морские свинки, свиньи, кролики, приматы), в том числе и у человека; все они имеют в крови высокий уровень БПЭХ. У животных, которые в крови имеют следовые количества этого белка (мыши, крысы, собаки), ЭХ не переходят из ЛПВП в ЛПНП и транспорт полиеновых ЖК в клетку проходит без участия апоВ-100 рецепторов. У этих животных ЭХ из ЛПВП переходят не в ЛПНП, а в ЛПОНП (апоА-1 + апоЕ БЛК); клетки активно поглощают ЛПОВП путем апоЕ/А-1 эндоцитоза. Активный транспорт полиеновых ЖК через апоЕ/А-1 рецепторы филогенетически более поздний и более резистентный к эндогенной блокаде. Мы полагаем, транспорт в клетки полиеновых ЖК путем апоВ-100 эндоцитоза обуславливает чувствительность животных к атеросклерозу, а апоЕ/А-1 эндоцитоз резистентность к атеросклерозу (Рис. 3).
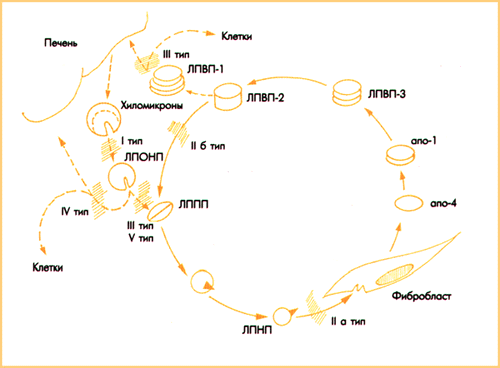
Рис. 3. Два варианта активного транспорта в клетки попиеноаых ЖК, которые определяют чувствительность (верх) и резистентность (низ) млекопитающих к атеросклерозу. Обозначения в тексте.
Атеросклероз, мы полагаем, является синдромом дефицита в клетках полиеновых ЖК. Симптомами атеросклероза являются ГЛП, гипертония, гиперагрегация тромбоцитов, гиперинсулинемия и диабет 11 типа, гиперкоагуляция, активация гуморального и клеточного звеньев иммунной системы и воспаление. Существенное различие транспорта в клетки ЖК дает основание дифференцировать два вида патологии - патологию транспорта полиеновых и патологию транспорта насыщенных ЖК. К первой относится атеросклероз, ко второй - эссенциальная гипертония.
Литература:
1. Титов В. Н. Вопр. мед, химии 1995. 1. стр. 3-8,
2. Титов В. Н. Биохимия 1996. 61. стр. 3-23.
3. Титов В. Н. Биохимия 1997. 62. стр. 3-19.
4. Титов В. Н. Успехи современной биологии 1997. 113.2, стр. 240-255.
5. Титов В. Н. Кардиология 1998. 38. 1. стр. 143-149.